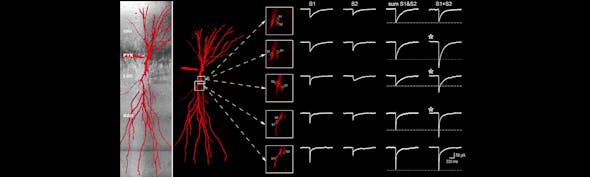
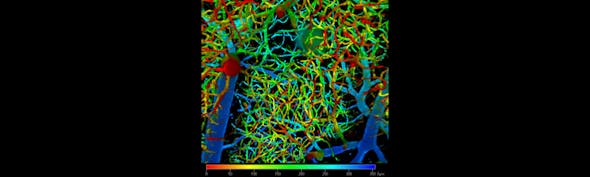
Challenges and Insights into Long-Term functional GCaMP imaging

by Florian Kämpf, Research Assistant in Armin Bahl’s Lab, University of Konstanz, Germany
Introduction
I'm absolutely fascinated by brains. The way neurons wire together with each other to create intricate networks without a clear blueprint is nothing short of astonishing. This awe turns my attention to GCaMP - a ground-breaking protein that allows me to peek into these circuits as they emerge from chaos.
But let's go back to the beginning. Since its debut in 2001 in mice, GCaMP proved itself by transforming Neuroscientists into part-time telepaths. It allowed us to monitor the activity of living neurons without harming the animals these neurons belonged to. GCaMP especially shined at the supervision of many neurons superseding the established method of electrophysiology. Combining this strength with the accessible and small brains of Drosophila, C. Elegans, and Zebrafish, GCaMP paved the way to a new era of circuit neuroscience which investigates how brain-wide neural circuits process information and influence behaviour.
For instance, the work of Green et al. (2017) et al. used GCaMP in Drosophila to unravel a neural circuit mechanism for angular integration, empowering little fruit flies to maintain and update their heading direction while walking. Meanwhile, Ngyuen et al. (2016) bridged the connection to behaviour by expressing GCaMP in freely moving C. Elegans. They identified clusters with neural activity correlated with locomotion behaviours including turning and forward/backward motion. But it doesn't stop there. Finally through the expression of GCaMP in larval zebrafish, Bahl and Engert (2020) illuminated a decision-making circuit, showcasing GCaMP's prowess in identifying circuit mechanisms responsible for higher cognitive functions.
In essence, GCaMP has paved a path to a ground-breaking era of neuroscience, one that meticulously investigates how single cells together process information and produce relevant behaviours. We now have a good overview of GCaMP and how it is currently used to identify circuits based on neuronal calcium dynamics. However, these approaches often look at circuits at very defined time points and overlook their part in a dynamic animal that constantly changes. Thus, I want to introduce you to the idea of long-term GCaMP imaging which allows us to observe how the circuits develop and how network-defining dynamics form over time. I will talk about the characteristics of GCaMP and some technical aspects to think about when you want to imagine an animal for 24h+. Moreover, I aspire to inspire you to observe your favourite circuit for an absurdly long time. So, prepare your lasers, secure your specimens, and ready your stimuli since we are about to jump right in.
Photobleaching
When we want to image animals with GCaMP for longer time scales an inevitable challenge we encounter is photobleaching. Photobleaching refers to the phenomenon during which fluorophores, like GCaMP, exponentially lose their ability to fluoresce over photon exposure. This is not only a weakening of the fluorescence of the affected molecules, but an irreversible destruction which we must avoid, since we want to image the same neurons again and again. As GCaMP molecules bleach, the signal-to-noise ratio deteriorates, making it increasingly difficult to discern genuine neuronal activity from background noise. This change in signal-to-noise ratio (SNR) makes the comparison of our observed circuit between different time points impossible.
Several approaches can be taken to tackle the photobleaching of GCaMP. On the imaging side, we can modify the exposure of the excitation light by reducing intensity and shorter exposure time. These parameters of course need to be counterbalanced with the loss in imaging quality. It's no use if you have no photobleaching but also no data to analyse. Additionally, GCaMP is continuously developed further, and while it is still not built for day-long recording, the new GCaMP8 version shows significantly lower photobleaching than previous versions (Looger, n.d.).
Moreover, other methods of microscopy intrinsically produce less photobleaching through their method to excite GCaMP. Two-photon microscopes for example use the energy of two photons to excite GCaMP compared to conventional confocal microscopes which only use one photon. Since the two-photon effect is only active in a defined focal plane, two-photon excitation produces photobleaching only in the plane of focus (Denk et al., 1990). For experimental optimization of long-term two-photon imaging, a trade-off between SNR and photobleaching must be found in terms of imaging power, since the ratio of power to photoemission increases slower than the ratio of power to photobleaching (Chen et al., 2002; Patterson & Piston, 2000). To minimize photobleaching in the focal plane during two-photon imaging, it has shown to be beneficial to have high concentrations of GCaMP. This means multiple inserts of the GCaMP sequence into the experimental animals' DNA (Hires et al., 2008; Yasuda et al., 2004).

In summary, although photobleaching is a significant challenge in long-term functional imaging, careful design and optimization of experiments as well as the utilization of advancements in microscopy and protein engineering make it a manageable problem.
Phototoxicity
Similar to photobleaching, phototoxicity is another phenomenon that we need to balance with SNR. This problem arises when the wavelength we use to excite our fluorophore, in our case GCaMP, induces toxic effects in the specimen we are trying to image. This problem is exacerbated by the fact that we plan to image the samples for long periods, which means that the tissue is repeatedly exposed to potentially toxic conditions.
For example, the energy absorbed during fluorescence can lead to the formation of reactive oxygen species, which can damage cellular structures such as the mitochondrion (Boudreau et al, 2016). Another source of phototoxicity may be the generation of heat during imaging at high laser powers, which may affect observed brain functions (Podgorski & Ranganathan, 2016). By altering the physiology of the neurons we observe, we are not able to measure the circuit of interest in its natural state, so we should try to minimize phototoxicity.
Reducing light intensity and exposure time, similar to the approaches used to combat photobleaching, can significantly reduce the risk of inducing phototoxic effects. Similarly, two-photon approaches can reduce toxicity during imaging by minimizing the area of effect in the focal plane. In addition, we must consider the welfare of experimental animals, as phototoxic effects most often represent permanent damage to DNA, can induce apoptotic responses, and generally do not promote healthy brain tissue (Tosheva et al.,2020).
Data Storage
Long-term imaging generates a lot of data that needs to be stored. This raises the question of whether to store raw data or pre-processed data. What format should the data be stored in? And finally, where to store the data, whether on a local hard drive or on a server system that allows access from the cloud. To give an example of how large a long-term imaging dataset would be, let's say we were imaging at 1Hz with a region of interest of 500 x 500 pixels for 24 hours at 16-bit integer bit depth. This would result in an uncompressed dataset of 43.2 gigabytes.
The Hierarchical Data Format (HDF) is an ideal data format for storing such large image datasets. Its accessibility to most programming languages, its ability to efficiently compress data, and its flexibility to store hierarchical data in custom organization make HDF an exceptionally versatile and scalable solution for the complex needs of neuroimaging data and its analysis.
Data Analysis
Once we have recorded the data we have to perform analysis. Software like Fiji (Schindelin et al., 2012) can help with first data explorations to get a feeling for the data and to get ideas about possible avenues of analysis. Further processing and analysis of the data should be performed in scripting languages like Python, Matlab, or R. This makes results easier to replicate and the analysis easier to share with collaborators and the public after publication.
One of the most standard methods for the analysis of GCaMP imaging data is the utilization of ∆F/F0 to put a cell/pixel fluorescence to a baseline F0. This baseline or base activity has to be chosen carefully based on the experimental paradigm and for example, can be a cell response to a neutral stimulus, or the average activity before an event of interest (Andreev, 2021).
In general, the ∆F/F0 can be calculated with:

Another important part of the GCaMP analysis is cell segmentation. Cell segmentation uses computer vision methods to detect the outlines of individual cells. Segmentation allows activity to be analysed at a single-cell level, taking advantage of the single-cell resolution of GCaMP imaging. This provides an alternative to pixel or voxel-based analysis. Segmentation algorithms operate on different principles such as pixel correlation (Zhang et al., 2014) or deep learning (Stringer et al., 2021).
Motion correction and drift
A major problem we have to consider, not only when imaging for long periods, but in general, is how our experimental animal moves while we are imaging it. In the ideal case, the animal does not move in its x, y, or z axis, but if we want to observe circuits, this is almost unavoidable. Of course, we can't anesthetize the experimental animal either, because that would introduce artifacts that would cause unnatural responses in the network dynamics.
Movements in x, y, and z can be caused by the animal responding with motor outputs to the displayed stimuli. Here, x and y shifts are easily corrected with motion correction algorithms (Giovannucci et al., 2019), allowing voxel/cell-based analysis. Z-shifts are more critical because they change the focal plane, meaning that we no longer observe the same cells. We can counteract these Z shifts either by including contingency plans in our imaging software that counteract Z shifts or by using wide-field imaging methods such as light sheet microscopy (Stelzer, 2015), which scans the whole brain several times per second.
Point scanning microscopes vs Plane illuminating microscopes
As we now already learned, long-term imaging is an endeavour of making compromises. This doesn't stop at the microscopes used to excite and detect the fluorophores of GCaMP. Point scanning microscopes, such as two-photon microscopes, excel in their ability to image deep within tissue with high spatial resolution and reduced photobleaching. However, its point-by-point scanning method can be slow, limiting its suitability for capturing fast, brain-wide events in multiple focal planes.

Plane-illuminating microscopes, like light sheet fluorescence microscopes, excel at imaging brain-wide activity in different planes through their mechanism of illuminating the plane of interest all at once. However, this brain-wide mode of capturing neural dynamics comes at a trade-off of reduced spatial resolution compared to point scanning microscopes, and its implementation requires the transparency of the experimental animal (like zebrafish or C. elegans) since illumination happens laterally to the brain.
Choosing between these different microscopy approaches for long-term imaging depends on the specific requirements of the study, including the need for depth penetration versus speed, the suspected localization of the investigated circuits, resolution versus field of view, and the sensitivity to photodamage of the tissue of interest.
Outlook
Now that we have overcome the many challenges of long-term imaging, I would like to ask the question: What do we learn when we look at a network for a long time? I'm most excited about observing the dynamic evolution of neural networks in real-time. How do they emerge and mature? Is there a recognizable precursor stage, or do functional circuits spontaneously emerge from previously dormant cell assemblies? These questions lie at the heart of neurodevelopment and neural plasticity and are not only of academic interest.
The observation of circuit development is highly relevant when applied to the study of neurological disorders. For example, long-term imaging of circuit formation in zebrafish models of autism provides a unique opportunity to determine when and how developmental trajectories diverge from wild-type patterns. Such insights could help elucidate the network pathology of autism and other disorders, and ultimately pave the way for new therapeutic strategies.
In summary, long-term functional GCaMP imaging holds great promise for neuroscience. Despite the challenges and complexities involved, the potential to gain deep insights into circuit formation and function justifies the effort. I look forward to future studies that focus more on neuron-based brain development.
About Florian
Florian completed his MSc with Armin Bahl Lab at the University of Konstanz, Germany; where he focused on 2-photon imaging techniques. He will shortly begin his PhD at the University of Cambridge in Gregory Jefferis lab.
References
Andreev, A. (2021) A biologist’s checklist for calcium imaging and optogenetic analysis. https://focalplane.biologists.....
Chen, T. et al. (2002) 'High-Order Photobleaching of Green Fluorescent Protein inside Live Cells in Two-Photon Excitation Microscopy,' Biochemical and Biophysical Research Communications, 291(5), pp. 1272–1275. https://doi.org/10.1006/bbrc.2....
Denk, W., Strickler, J.H. and Webb, W.W. (1990) 'Two-Photon laser scanning fluorescence microscopy,' Science, 248(4951), pp. 73–76. https://doi.org/10.1126/scienc....
Hires, S.A., Tian, L. and Looger, L.L. (2008) 'Reporting neural activity with genetically encoded calcium indicators,' Brain Cell Biology, 36(1–4), pp. 69–86. https://doi.org/10.1007/s11068....
Patterson, G.H. and Piston, D.W. (2000) 'Photobleaching in Two-Photon excitation microscopy,' Biophysical Journal, 78(4), pp. 2159–2162. https://doi.org/10.1016/s0006-...(00)76762-2.
Schindelin, J. et al. (2012) 'Fiji: an open-source platform for biological-image analysis,' Nature Methods, 9(7), pp. 676–682. https://doi.org/10.1038/nmeth.....
Yasuda, R. et al. (2004) 'Imaging calcium concentration dynamics in small neuronal compartments,' Science’s STKE, 2004(219). https://doi.org/10.1126/stke.2....